
The Origin of Peoples
| by | Trevor Major, M.Sc., M.A. |
As we look among the peoples of the world—from the Inuit to the !Kung, from the Norwegian to the Greek, and from the Indian to the Tutsi—we see a mind-boggling array of skin color, hair type, stature, and facial features. On top of all that physical diversity, we must add differences in culture and language. With technological advances, humans have lived (if only for a short time) at the South Pole, on the peaks of the Himalayas, and beyond Earth itself. Even before the advent of modern science, we have occupied the remotest islands, the driest deserts, and the coldest steppes. It is difficult to imagine any other creature that has been so successful at colonizing so many different parts of this planet (we’ll give the cockroach its due!).
For all these differences, we constitute a single, biological species. Men and women with roots in different continents meet, marry, and have healthy families. This unity frustrates any attempt to parcel the world’s populations into distinct subspecies or races. We perceive great diversity because our brain is so cleverly designed to detect patterns and distinguish among individuals of our own kind. Such heightened perception of the human form is something we cannot ignore, and shapes a host of psychological responses such as physical attraction and group identity. Still, at the biological level, this variation reflects minute differences in our genetic code. We see a few of these in our physical appearance, but find many more only at the cellular or molecular level. One person may have resistance to a particular disease, while another is able to digest milk as an adult. Whether on the inside or outside, the combination of many subtle differences makes you and me stand out as individuals within a group, and our similarities identify us with humanity as a whole.
How did these differences arise? Like Rudyard Kipling’s Just So stories, we could spin all sorts of tales to explain why different peoples are the way they are. We could tell a story about how the Scandinavians became tall, and another story about how they became light-skinned. The goal for this traditional Darwinian approach is to answer the following question: How does a particular trait enhance survival value, or enable the production of more offspring? One anthropology textbook emphasizes the “pervasiveness of adaptation in the microevolution (small-scale differentiation) of man” (Keesing and Keesing, 1971, p. 51). As we will see, this turns out to be more of a hope than a claim based on evidence.
There is the assumption, also, that we need a lot of time to explain human variation because evolution works at a steady, snail’s pace. Charles Darwin took this as a matter of principle, but not all evolutionists agree. A few dissenters, citing examples from the fossil record, believe that species arise during brief moments of intense change, rather than by slow accumulation of new features (e.g., Eldredge, 1985, pp. 21-22). So, too, within human populations, distinct groups might arise during significant natural or cultural events. In addition, more evolutionists are expressing concern about the “molecular clock.” This was supposed to represent the rate at which genetic differences have accumulated in two related species. However, the calculation depends on knowing the date of the presumed common ancestor. Not everyone may agree on this date, or even on whether the two species are closely related. In any case, evolutionists assume that humans have diverged from each other at about the same rate we diverged from chimpanzees—our supposed closest relative. However, a closer look at families of known lineage has revealed mutation rates that are almost twenty times higher than previous estimates (Gibbons, 1998). The upshot is this: we cannot trust the Darwinists’ intuitions on the time it would take to produce the differences we see in human populations. The rate may be neither slow, nor steady.
For the moment, I would like to set aside the question of time (but see my sidebar article), and focus on the biological bases for some of the differences that have arisen among our kind.
IN LIVING COLOR
The difference we tend to notice most is coloration, which depends almost entirely on the relative abundance of melanin. This is a pigment of the hair, skin, and irises. It seems to play a role in protecting the skin from harmful ultraviolet rays. Exposure to the Sun increases melanin, causing that tanning effect so prized by light-skinned Westerners. At first glance, it looks as if the inhabitants of equatorial regions, where sunlight bears down with the greatest intensity, would have the most melanin. After all, sub-Saharan Africans, and Australian Aborigines, have more melanin than northern Europeans.
Around 1913, Charles Davenport suggested that humans carried two genes for color, and that each gene consisted of “black” or “white” alleles (one allele from the mother, and one from the father, for each gene). Hence, our coloration depends on the number of black and white alleles we received from our parents. Davenport noted correctly that children inherit these genes independently of other characteristics, such as straight versus curly hair. This explains why albino Papuans look different from albino Scots.
As usual, the advance of science has revealed a far more complicated story. Geneticists now believe that almost half a dozen genes have a significant effect on pigmentation (Wills, 1994, pp. 78-79). These genes reside in the nucleus of every cell in our body, along with copies of all the other genes we inherited from our parents. However, color genes express themselves in only one place—the melanocytes. These are specialized skin cells that have a monopoly on melanin production. Each melanocyte is an incredibly complex chemical factory, transforming raw materials into granules of melanin, which it delivers to neighboring cells.
Also, there is more to the making of skin color than turning genes on or off to make black, white, and a couple of shades in between. We all possess the essential ingredients for making melanin; all of us could be black or brown (the only exceptions are albinos, whose bodies make no melanin at all). Actual coloration varies according to the pigment package delivered by the melanocytes. The end product depends not only on slight genetic differences, but also on environmental stimuli (such as exposure to strong ultraviolet radiation).
The story does not end there. Skin also includes keratin—a fibrous protein that contributes to the toughness of the skin, and which grows to form nails and hair. Because this substance has a relatively high concentration of sulfur, it adds a yellow hue to our palette of skin colors. Asians (especially from the Far East) happen to have an extra thick layer of keratin which, when combined with melanin, contributes to the yellow-brown color of their skin.
The science of genetics helps us understand how small changes can account for the rainbow of human coloration. Truly, when we consider the magnitude of these differences at the genetic level, our obsession with skin color seems blown out of proportion.
NATURAL SELECTION AND HUMAN VARIATION
We know that there are variations in features such as skin color. Why, or how, did these variations arise? As noted earlier, a knee-jerk response is to invoke natural selection, but there are a few good Darwinian tales.
For instance, around 40% of the people in equatorial Africa carry an abnormal hemoglobin gene that deforms red blood cells into a crescent or sickle shape. Anyone who carries this trait, plus a normal copy of the gene, may appear to have the best of both worlds. For a start, the normal gene is dominant, and so counteracts the recessive mutated gene. Then, if malarial parasites invade the red blood cells, there is a tendency for the cells to deform and die, along with their unwanted guests. Unfortunately, people who have two copies of the abnormal gene develop sickle-cell anemia, and will die an early death unless they have access to good medical treatment. Finally, anyone not “lucky” enough to inherit the abnormal gene has no anemia, but no immunity from malaria either.
Of course, the picture is not all rosy for the people who carry just one copy of the sickle-cell gene. If they marry another carrier, some of the children could inherit two bad copies, and suffer from sickle-cell disease (see diagram below). With this in mind, it is callous to speak of the sickle-cell trait as a “good” or “beneficial” mutation. Nonetheless, the trait persists because the threat of death from malaria appears to outweigh the threat of death from sickle-cell anemia. In this instance, nature may have preserved a particular trait because it confers a survival advantage.
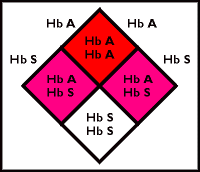 |
Sickle-cell genetics: In this example, two parents each have a normal (Hb A) and an abnormal (Hb S) hemoglobin allele. There is a 1 in 4 chance that a child will have normal hemoglobin (Hb A/Hb A), a 1 in 2 chance that a child will be a carrier for the sickle-cell trait (Hb A/Hb S), and a 1 in 4 chance that a child will have sickle-cell anemia (Hb S/Hb S).
|
For most variations that give human populations their distinctive characteristics, it is difficult to know what forces of selection have been at work. For instance, scientists used to think that the Pygmy people of southern Africa were short because food was scarce. Further studies show normal levels of growth hormone, but reveal a genetic defect that prevents their bodies from using the hormone to its fullest extent (Fackelmann, 1989). But the question is this: Did nature select this mutation because it offered survival advantages, or did this characteristic arise as a result of random variation?
The answer is not so obvious, because we know so many exceptions to the rules of natural selection. Take the Japanese, for instance. Their teenagers are considerably taller than their grandparents ever were. The difference is a matter of improving diet, not genetics. For hundreds of years, the people of Japan have survived without nature’s selecting mutations for smaller stature. So how do we know that a scarce food supply was responsible for the survival of growth-limiting changes in the Pygmy?
The list of just-so stories is endless. Why are the Inuit relatively short and bulky? Because this helps them retain heat. Why are some groups in Africa relatively tall and slender? Because this helps them lose heat. In each case, we could list a dozen exceptions. What about those tall peoples who have survived quite well in cold areas, like the Dutch? And what about those short peoples who have done just fine in hot areas, like the Pygmies?
If Africans have less hair to keep them cooler, as some have suggested (Folger, 1993), then how have Asians done so well in cold climates with relatively little body hair? Asians also have an epicanthic fold—an extra layer of skin on the upper eyelid. We could spin a story about their eyes adapting to the winds of the Mongolian steppes, or the bright glare of snow. Even so, is this enough? Are variations in the structure of the eyelid a matter of life and death? Were individuals who had this epicanthic fold much more likely to survive than those who lacked it?
Similar questions confront the origins of skin color. Precisely how has natural selection worked to preserve dark and light skin coloring? The traditional explanation makes what seems to be a sensible link between the strong sunlight of the tropics, and the protective powers of melanin. Natural selection, so the argument goes, has favored the survival of dark-skinned people in equatorial areas. If light-skinned people lived in the tropics, they would suffer from higher rates of skin cancer. Then what prevented Africans from migrating to higher latitudes? The answer, we are told, lies in vitamin D. To make this important substance, humans need exposure to ultraviolet light. If people in higher latitudes were too dark, their skin would not be able to make enough vitamin D. A shortage of vitamin D results in rickets, which has a severe effect on bone development. So everything works out perfectly: light people get a little melanin to avoid rickets; and dark people get a lot of melanin to avoid skin cancer. Whatever the explanation, many researchers remain convinced that some sort of evolutionary process must be responsible for lighter and darker strains of humans (see Wills, 1994, p. 80).
The story seems less plausible, however, when we try to imagine how selection might have worked. For instance, skin cancer is deadly; it is something that afflicts lighter-skinned people who spend much time in strong sunlight. People of European ancestry living in the sunny climes of Australia, New Zealand, and Hawaii suffer the highest rates of skin cancer in the world. As we look back in history, however, the danger of dying from basal cell carcinomas and melanomas hardly would compare to the vagaries of childhood diseases, plagues, strife, starvation, and natural hazards. It is hard to imagine that in a mixed population of light-and dark-skinned people living near the tropics, evolution selected the traits for dark skin because cancer gradually eliminated their lighter-skinned neighbors.
Unlike the skin cancer scenario, the ability to produce sufficient vitamin D is a definite survival advantage. However, exposure to the Sun is not an absolute requirement. Oils from cod, halibut, sardines, salmon, and mackerel provide a rich source of vitamin D (Sackheim and Lehman, 1994, p. 516). Not surprisingly, such fish figure prominently in the diets of Scandinavians and the Inuit. With the right foods, they are able to overcome a disadvantage of living in areas where the Sun is weaker, and in which the cold climate dictates many layers of protective clothing.
Still, this does not explain why Africans remained in tropical zones. They could have moved northward, and endured doses of cod liver oil as much as any European child. Today, thanks to vitamin supplements, people of African descent survive in England and Canada without a high incidence of rickets. When we look to the original population of the Americas, the story blurs altogether. People of brownish complexion live across every climatic zone, from Alaska in the north to Tierra del Fuego in the south. Apparently, no mechanism has been at work to sort skin color by latitude.
There are many other problems with the climatic theory of skin color (Diamond, 1992, pp. 114-117), and still, we have barely touched the rich storehouse of human variety. Perhaps apparently neutral characteristics will turn out to have some survival advantage (Patterson, 1978, p. 70). For example, researchers have found a correlation between ABO blood groups and resistance (or susceptibility) to different diseases. Further, blood groups seem to have a strong geographic distribution. We may discover that a particular blood type became concentrated in a region where it offered a slightly better chance of survival. On this point, however, all we have so far is another Kiplingesque story. No doubt, natural selection has had some impact on human history, but it seems largely inadequate to explain a good portion of the variations that exist between different human populations.
THE MAKING OF A PEOPLE
If natural selection has played a minor role in human history, then how do we explain the range of observed features? One possible mechanism is a phenomenon known as the “founder effect.” We see this most often in small, isolated communities that have an unusually high incidence of rare, inherited disorders (Diamond, 1988, p. 12). After some genealogical detective work, medical researchers are able to trace their patients’ ancestries to a single couple or a small group of close relatives—the founders. This seems to be the case with French Canadians, particularly those of eastern Quebec, whose ancestors emigrated from the Perche region of France in the 17th century. Small pioneering groups, together with early marriages, large families, and isolation, have created a pronounced founder effect. One study found that only 15% of the settlers contributed 90% of the genetic characteristics in people suffering from one or more of five genetic disorders (Heyer and Tremblay, 1995).
 |
Pioneers in Chicoutimi (c. 1886), which is now the modern administrative center of Saguenay-Lac-St-Jean. This part of Quebec was settled by a few, closely related families. Today, 9 or 10 rare genetic diseases are relatively common among the people of the region.
|
It is only natural that much of our information on founder effects should come from the study of debilitating, and often fatal, diseases. If medical researchers can pin the problem to a faulty gene, then this may suggest a treatment or cure. Also, genetic testing can tell prospective parents whether they will pass these mutations on to their children. If the effects of the disease will come later in life, people may want to start certain medical treatments, or make changes in diets, that will ease or delay the worst symptoms.
However, the record books include a few cases not related directly to diseases. In a now classic study, H. Bentley Glass (1953) found that the Dunkers—a community of German Baptist Brethren in Pennsylvania’s Franklin County—are, in most respects, very similar to other people of European descent. Their religious customs require them to dress a certain way, and marry within the community, but otherwise their physical appearance is not unusual. Although there have been some outside marriages, most of the surviving members are descended from fifty families that emigrated from Germany in the early 1700s. Glass found that the frequencies of blood types and other genetic traits among the Dunkers differ from the frequencies of these features among U.S. and German populations. It seems unlikely that any selective forces were in operation to favor the survival of Dunkers with blood group A, for instance. Therefore, Glass concluded, the founding population of Dunkers included, purely by chance, an unusually high proportion of people with blood group A.
The founder effect itself is part of a broader concept known as genetic drift, which occurs anytime the frequency of a genetic trait changes within a population. If, in the case of the founder effect, the emigrating group carries a set of unique or rare traits, then those traits will be that much harder to find among the people who stay behind. In other words, there will be a drift away from those characteristics.
In some cases, a highly prolific individual or family may skew the genes of a relatively diverse population, and this may occur in combination with some other form of genetic drift, such as the founder effect. For example, groups of Ashkenazic Jews moved eastward out of Germany in the 17th century, and were isolated culturally from the surrounding population. Several rare inherited disorders, such as Tay-Sachs disease, afflict this group at high rates. Evolutionists have thought this to be a sign of natural selection at work. Perhaps the population hung on to these genes because they offered some survival advantage, such as resistance to tuberculosis and other maladies of the crowded ghettos in which they lived (Diamond, 1991). However, Neil Risch believes otherwise, at least in the case of idiopathic torsion dystonia, which occurs at a rate of one in three thousand among the Ashkenazim today (Glausiusz, 1995). First, migration patterns favor genetic drift via the founder effect in these people. And second, historical records show that wealthier couples had more children. If a mutation arose in one of these families, as Risch infers from the genetic data, then it could become more frequent in later generations. This is a matter of misfortune, not adaptation.
Of all the forms of genetic drift, population bottlenecks are the most dramatic. Typically, these occur when wars, natural disasters, epidemics, and other catastrophes wipe out all but a small remnant of the original population. For instance, a flood could drown an entire tribe, except for a fortunate few in a remote village. These survivors would bequeath their genetic characteristics to subsequent generations. If there were a high degree of relatedness among the survivors, then their descendents may appear quite distinct from neighboring peoples. Of course, the Bible shows the Flood of Noah to be the greatest bottleneck of all time. According to the Genesis account, all of us must trace our ancestry to Noah’s three sons and their wives.
Finally, another piece of the puzzle may be mate selection. We are quick to point out the ways in which we differ from our spouse, and we see a positive side to that. “Opposites attract,” so the saying goes, but the Beach Boys knew better. “I seen all kinds of girls,” the Californian band harmonized, but “I wish they all could be California girls.” Underneath the superficial differences lie the grand similarities. Not always, but more often than not, we marry someone who grew up nearby, speaks the same language, and belongs to the same cultural, religious, social, and political group (Diamond, 1992, pp. 99-109). The result is a barrier, obvious or otherwise, that may exist between two neighboring peoples, or even between groups who live cheek-by-jowl.
THE BIBLICAL VIEW
Evolutionists may argue that an explanation for human diversity simply is unavailable to anyone who adopts a literal interpretation of the Bible. They may reason that creationists have no access to any mechanism that would cause change, because this means accepting evolution. This is a common misunderstanding. Creationists object, not to microevolution, but to macroevolution. One works by natural selection acting on mutations to create limited variation; the other assumes unlimited variation. One seems to work; the other is highly problematic. For our present purposes, we need account only for variation on a small scale, and within a single species at that. There is no reason to eliminate adaptation out of hand, especially as it seems to work in cases like sickle-cell anemia.
Further, many evolutionists imagine an entirely Darwinian plot. This may seem to threaten the biblical view on the grounds of time, assuming that adaptation implies a slow, gradual process. Not everyone agrees on this tempo of change and, certainly, genetic studies are revealing ample non-Darwinian strategies.
The key biblical event must be the confusion of tongues at the Tower of Babel (Genesis 11). Up to this point, as far as we can tell, three lines of descent were living in close proximity, and then a miracle occurred. God gave them different languages so they could not work together on the Tower (11:7). They could have dug their heels into the rich soil of the Fertile Crescent, and trained a few good translators, but God “scattered them abroad” (11:8).
We cannot be sure on what basis the partitioning occurred. In the Table of Nations (Genesis 10), each line of descent appears by family and language, according to their lands and nations (10:5,20,31). It seems most likely, therefore, that the division occurred by the principal family units present at the time of the confusion and dispersion. This corresponds to the time of Peleg, in whose days “the earth was divided” (10:25). It is at this point that the mechanisms described earlier must come into full force. If the human population scattered over the face of the Earth, then there was a sudden outpouring of founding groups. Each extended family, isolated from others by language, would carry its own set of genes into the world. From these groups, and within these groups, developed the peoples of the world.
REFERENCES
Diamond, Jared (1988), “Founding Fathers and Mothers,” Natural History, 97[6]:10-15, June.
Diamond, Jared (1991), “Curse and Blessing of the Ghetto,” Discover, 12[3]:60-61, March.
Diamond, Jared (1992), The Third Chimpanzee (New York: HarperCollins).
Eldredge, Niles (1985), Time Frames: The Evolution of Punctuated Equilibria (Princeton, NJ: Princeton University Press).
Fackelmann, K.A. (1989), “Pygmy Paradox Prompts a Short Answer,” Science News, 136[2]:22, July 8.
Folger, Tim (1993), “The Naked and the Bipedal,” Discover, 14[1]:34-35, November.
Gibbons, Ann (1998), “Calibrating the Mitochondrial Clock,” Science, 279:28-29, January 2.
Glass, H. Bentley (1953), “The Genetics of the Dunkers,” Scientific American, August. Reprinted in Human Variation and Origins (San Francisco, CA: W.H. Freeman), pp. 200-204.
Glausiusz, Josie (1995), “Unfortunate Drift,” Discover, 16[6]:34-35, June.
Heyer, E. and M. Tremblay (1995), “Variability of the Genetic Contribution of Quebec Population Founders Associated to Some Deleterious Genes,” American Journal of Human Genetics, 56[4]:970-978.
Keesing, Roger M. and Felix M. Keesing (1971), New Perspectives in Cultural Anthropology (New York: Holt, Rinehart, and Winston).
Patterson, Colin (1978), Evolution (London: British Museum/Cornell University Press).
Sackheim, George I. and Dennis D. Lehman (1994), Chemistry for the Health Sciences (New York: Macmillan).
Wills, Christopher (1994), “The Skin We’re In,” Discover, 15[11]:76-81, November.
No comments:
Post a Comment